1. Introduction
Fibers are thread-like structures with continuous or discrete phases of filaments. The predominant advantage of natural fiber is biodegradability, lightweight, and less density. Natural fibers account for innumerable benefits of sustainability and eco-friendliness. The concern about natural fibers research has increased recently due to the need for an alternative towards synthetic fiber usage. Some cost-effective reinforcement materials from the natural fibers pre-owned as an alternative to synthetic fibers are flax, jute, ramie, hemp, banana, etc. (Chabba et al., 2005). The natural fiber application has been inclined tremendously since the last decade. Jain et al. (2016) reported that the dislocation of famous designers and renowned fashion brands was searching for eco-friendly fibers to make sustainable products for clothing and other apparel. Natural fibers account for cost-effectiveness, manageable processing, and renewable resources. The natural fibers can be classified as plant-based, animal-based, and mineral-based. The plant-based fibers can be further categorized as bast fibers, leaf fibers, wood fibers, fruit fibers, seed fibers, straw fibers, and grass fibers. Our research paper consolidates the degumming of plant-based natural fibers for textile applications.
1.1. Plant-based fibers
Seed fibers: Among the natural fibers, Cotton fiber is a highly explored natural fiber. The source of cotton fiber is plant seeds, predominantly known as seed fibers. Cotton fiber belongs to the genus Gossypium. Cotton fibers are enriched with cellulose. The 90% composition of Cotton fiber possesses cellulose. The characteristic features of cotton fiber are absorbent, lightweight, and soft. The major applications of cotton fiber are textiles, woven fabrics, fishing nets, and so on. Other seed-based fibers are Pine-cone, Kapok, Loofah, and rapeseed (Pinheiro et al., 2020).
Bast fibers: The members of bast fibers are Flax, Jute, Ramie, Kenaf, Hemp, Mesta, and Roselle. Hemp fibers are identified to be one of the strongest members of bast fibers. Hemp fibers are derived from the species Cannabis. The composition of cellulose in hemp fiber is about 70%. Ramie fibers are powerful bast fibers with long durability and higher cellulose content. Ramie fibers are also known as china-grass, grass linen, grasscloth, and china linen. The durability and strength of ramie fibers promote its application in industrial sewing thread, packaging materials, fishing nets, and filter cloths.
Similarly, Jute fibers, commonly known as lignocellulosic bast fibers, belong to the genus Corchorus. Kenaf fibers are derived from the bast of Hibiscus comes under the family Malvaceae. The features of kenaf fibers are flexural strength, lightweight, ignition, biodegradability, and eco-friendliness (Jain et al., 2016).
Grass fibers: The grass fibers are Bamboo, Bagasse, Corn, Sabai, and Canary. The bamboo fibers are derived from Bamboo plants that belong to the family Bambusoideae. The bamboo fiber has enormous micro-gaps, which are responsible for higher absorption quality. The features of bamboo fibers are antimicrobial, hygroscopic, and resistant against UV light.
Fruit fibers: Coir fiber and Oil-Palm fiber comes under fruit fibers. Coconut or coir fibers are obtained from the husk of the coconut. The coir fiber belongs to the family Cocus nucifera. The properties of coir fibers are durable and cost-effective. Oil-palm fibers are also known as lignocellulosic fibers, comes under the family Arecaceae. The source of oil-palm fibers is fruit mesocarp and empty fruit bunch of oil-palm tree.
Leaf fibers: The leaf fibers are Sisal, Banana, Abaaca, Pineapple, Henequen, and Agave. Banana fiber (lignocellulosic fiber) is generated by the pseudostem, which has significant mechanical properties (Drahansky et al., 2016). Banana pseudostem seems to be clustered, cylindrical solicitation of leaf stalk bases. The banana pseudostem can be implemented for making marine rope, coffee, tea bags, filter cloths, and light-density woven fabrics. (2010) mentioned the consideration towards banana pseudostem in textile, paper, and composite material for structural reinforcement. The pseudostem exhibits antimicrobial properties (Bello et al., 2018). Sisal fibers are the cellulosic fibers derived from the Agave sisalana. The sisal fibers are perennial succulents with major applications in rope-making, paper, cloth, geotextiles, dartboards, footwear, hats, and bags. Sisal fiber belongs to the family Asparagaceae.
Wood fibers: Wood fibers can be classified as softwood and hardwood fibers. The major components of wood fibers are lignin, cellulose, hemicellulose, and extractives. Both kinds of wood fibers play a major role in paper manufacturing. The properties of natural fibers such as banana, coir, cotton, flax, hemp, jute, kenaf, palm, pineapple, ramie, sisal, bamboo, and dates are listed in the following Table 1 (Paluvai et al., 2014) (Sreenivas et al., 2020).
Table 1. Properties of Natural fibers.
| Fiber type | Density (Kg/m3) | Cellulose (wt %) | Lignin (wt %) | Tensile strength (Mpa) | Young modulus (Gpa) | Elongation (%) | Moisture content (wt %) |
|---|---|---|---|---|---|---|---|
| Banana | 1,350 | 63–64 | 15 | 529–914 | 27–32 | 1–3 | 13 |
| Coir | 1,150 | 32–45 | 40–55 | 140–593 | 4–6 | 25–30 | 8 |
| Cotton | 1,600 | 85–90 | 0.4–1 | 287–597 | 5.5–12.6 | 7–8 | 7.85–8.5 |
| Flax | 1,500 | 71 | 2.2 | 500–1500 | 27.6 | 2.7–3.2 | 8–12 |
| Hemp | 1,480 | 70–74 | 3.7–5.7 | 550–900 | 30–70 | 1.6–1.8 | 6.2–12 |
| Jute | 1,460 | 61.1–71.5 | 12–13 | 393–773 | 26.5 | 1.5–1.8 | 12.5–13.7 |
| Kenaf | 1,400 | 45–57 | 8–13 | 930 | 53 | ||
| Palm | 0.7–1.5 | 42.7–65 | 13.2–25 | 50–400 | 0.57–9 | 2.5–18 | |
| Pineapple | 1,440 | 70–82 | 5–12.7 | 640 | 2.4 | 11.8 | |
| Ramie | 1,500 | 68.6–76.2 | 0.6–0.7 | 400–938 | 61.4–128 | 3.6–3.8 | 7.5–17 |
| Sisal | 1,450 | 66–78 | 10–14 | 511–680 | 9.4–22 | 2.0–4 | 10–22 |
| Bamboo | 910 | 503 | 35.91 | 1.4 | |||
| Date | 990 | 309 | 11.32 | 2.73 |
1.2. Need for fiber degumming
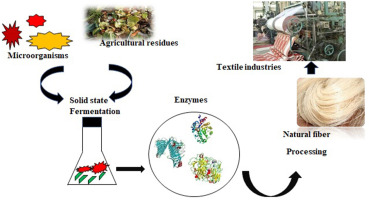
Industrialization of natural fibers necessitates removing non-cellulosic content of the threads, which degumming fibers can accomplish. Natural fibers, namely jute, ramie, kenaf, and hemp, have significantly experimented for fiber degumming application and production of value-added products (Chiliveri et al., 2016). Banana fibers are found to be less explored natural fibers in the degumming application. Degumming eliminates the viscous or sticky consistency from the cellulosic part of plant fibers to increase textile manufacturing industries. The techniques of degumming methods are acid, alkali, steam explosion, ultrasonic, microwave, bacteria, fungus, and enzyme (Shen et al., 2015). The microbial degumming has declined the chemical and energy consumption of natural fiber.
1.3. Microbial degumming
The enzymatic degumming process is an eco-friendly method as it reduces fiber damage without altering the properties of cellulosic fibers (Patidar et al., 2018). The pectinase and xylanase were identified to be adequate enzymes for fiber extraction and degumming process (Tibolla et al., 2014) (Jacob et al., 2008). In microbial degumming, fibers will be autoclaved for 20 min. After sterilization, degumming temperature, degumming time, and enzymatic dosage for the degumming of natural fibers will be optimized. The microbial pectinase is noteworthy in fiber processing as plant cambium composes 40% pectin in dry weight (Bruhlmann et al., 1994). The hemicellulose and lignin removal from the fiber surface results in single fiber formation, which will be further processed for textile applications(Angelini et al., 2015). Kohli et al. (2019) confirmed the eco-friendliness and energy conservation by adopting the pectinolytic enzymes in plant fibers degumming. (2013) suggested the highlights of utilizing the nanoparticle-based pectin degrading enzymes to reduce sugars and further application in the degumming of ramie fiber.
2. Pectinase
Pectinases can be of different forms based on their activity on the structure of polygalacturonate. Enzymatic activity differs concerning the degree of methylation. The enzymatic and specific activity of pectinase relies on the methylation degree. The chrysanthemum family was identified to be the first structure of pectinase. β-helix, distinct β-sheets from β strand turns are the structural topology of the pectinase enzyme (Henrissat et al., 1995). The active sites are His223 and Asp201, which act as a donator of proton and nucleophile, respectively. His223 provokes proton transfer, glycosides bond breakage, and generation of the covalent bond. Simultaneously, the opponent residue restores the enzyme active site by locating the H2O molecule to rescue the second product (Palanivelu, 2006). Fig. 1 illustrates the 3-dimensional image of the pectinase enzyme.
 Fig. 1
Fig. 1Some of the prevailing microorganisms involved in pectinase production are Pseudomonas, Xanthomonas, Erwinia, Actinomycetes, and Streptomycetes and fungal sources such as Aspergillus niger, Aspergillus versicolor, Aspergillus flavus, Fusarium oxysporum, Rhizopus stolonifer, Mucor racemous, Mucor hiemalis, Penicillium jenseni, Penicillium citrinum and Trichoderm aviride (Priya and Sashi, 2014). The convenience of microbes results in the decline of fiber damage, energy consumption, and non-toxic effluents. Banik et al. (2008) put forward the exploitation of pectinolytic microbes in the piling of jute fibers and their ability to enhance yarn quality. The propensity of pectinases in the breakdown of complex molecules, retting, and degumming of fiber crops are remarkable (Chiliveri et al., 2016).
2.1. Production of pectinase by fermentation
In contrast to submerged fermentation, the supremacy of low contamination, high yield, utilization of agricultural residue as substrates, simplicity of processing, and lower energy requirements were the convenience of solid-state fermentation (Selvakumar et al., 1998). The solid substrate fermentation process is more than 5.87 times advantageous than other processes (Kapilan and Arasaratnam, 2011). Investigation on the substrates suggests sugar cane bagasse, wastes of tea, cassava, apple, oil mill, banana, corn, wheat (straw, bran, husk, flour), coconut coir pith, rice (straw, bran), sawdust, the pulp of sorghum and sugar beet, cake of rapeseed, coconut oil, etc. for the fermentation. The Pectinase production using various species and substrate are listed in Table 2.
Table 2. Pectinase production using various species and substrate.
| Species | Substrate | Type of Fermentation | pH | Temperature | References |
|---|---|---|---|---|---|
| Aspergillus niger LFP-1 | Pomelo peel | solid substrate fermentation | pH 4.5 | (30 °C) | Darah et al. (2015) |
| Bacillus subtilis |
Hazelnut shell Hydrolysate |
submerged fermentation | pH 7.0, | 30 °C | Uzuner and Cekmecelioglu (2015) |
|
Bacillus tequilensis SV11-UV37 |
wheat bran | solid substrate fermentation | pH 6 | 37 °C, | Chiliveri et al. (2016) |
| Aspergillus oryzae | citrus waste and sugarcane bagasse | Solid substrate fermentation | pH 4 | 47 °C | Biz et al. (2016) |
| Rhizopus sp. C4 | Peel of orange | Solid substrate fermentation | pH4-5 | 30 °C | Handa et al. (2016) |
| Bacillus subtilis SAV-21 | orange peel and coconut fiber (4:1) | Solid substrate fermentation | pH 4.0 | 35 °C | Kaur and Gupta (2017) |
| Aspergillus fumigatus R6 | rice bran | Solid substrate fermentation | pH4-5 | 33 °C | Wong et al. (2017) |
| Aspergillus niger | orange pomace | solid-state fermentation. | pH 4.0 | 45–55 °C | Mahmoodi et al. (2017) |
| Bacillus sp. Y1 | wheat bran | Solid substrate fermentation | pH 8.2 | 34 °C | Guo et al. (2019) |
| Aspergillus tamarii. | Chikoo peels and pulp | submerged fermentation | pH (3–6) | (30–60 °C), | Munir et al. (2019) |
| Aspergillus niger | Biserrata leaves | Solid substrate fermentation | pH 4.0 | 50 oC | Pagarra et al. (2019) |
| Aspergillus niger | Nephroleis biserrata | Solid substrate fermentation | pH 4.0 | 50 oC | Pagarra and Rahman (2019) |
| Aspergillus sp.VTMS | Coffee Pulp | Solid substrate fermentation | pH (3–5) | 30 oC | Hidayah et al. (2020) |
| B. licheniformis KIBGE-IB3 | Date fruit waste | Solid substrate fermentation | pH 7.0 | 37 oC | Aslam et al. (2020) |
| Aspergillus niger | Pineapple peel pectin | Solid substrate fermentation | pH 5.0 | 40 oC | Ajayi et al. (2021) |
2.2. Application of pectinases in degumming of fiber
Textile application of natural fibers prerequisite the degumming process, which detaches the non-cellulosic material. The enzymatic degumming overwhelms the pitfall of alternate methods (Baracat et al., 1991). Kashyap et al. (2001) reported that Bacillus enzyme treatment was used in the retting of Mistumatabast fibers, yielding high profitability compared to other methods. Commercial alkaline pectate lyase is more capable of flax retting with high tenacity and low structural damage (Jain et al., 2016). Kaur et al. (2016) disclosed Bacillus pumilus BK2 as the valuable cotton scouring source. Alkaline pectinase has great scrutiny in the textile industry to replace water loss, energy demand, and toxic chemicals. They play a major role in degumming bast fibers, ramie fibers, flax fibers, and other eco-friendly fibers. The synthesis of pectinases from various species under different operating conditions and their advantages are listed in Table 3.
Table 3. Degumming of natural fibers by Pectinases produced from different species.
| Species | Enzyme | Application | Operating Conditions | Observation | Problems | References |
|---|---|---|---|---|---|---|
| Pseudozyma sp. SPJ (Yeast) | Pectinase | Flax fiber degumming | 12 h incubation | Weight loss of 11 ± 1.2% | No studies on galacturonide release | Sharma et al. (2011) |
| Bacillus megaterium AK2 (Bacteria) | Pectate lyase | Ramie fiber degumming | 24–48 h processing time | Nanoparticle supplemented pectate lyase improves weight loss% | Prolonged incubation | Mukhopadhyay et al. (2013) |
| Bacillus tequilensis SV11-UV37 (Bacteria) | Pectate lyase, Poly-galacturonase | degumming of kenaf, sun hemp fiber |
300 U at 50 °C in 11 h +NaoH treatment |
Weight loss, galacturonide acid was higher with combined methods | Usage of chemicals | Chiliveri et al. (2016) |
|
Aspergillus niger Aspergillus fumigatus (Fungus) |
Pectinase Enzyme | Banana Fiber extraction | 30 °C incubation for three days | Higher pectinolytic activity under solid-state, the tensile strength of fiber improved | No studies on weight loss | Sarma and Deka (2016) |
|
Fusarium oxysporum MTCC 1755 (Fungi) |
Pectin lyase | Hemp fibers |
Glycine NaOH treatment + 24 h incubation |
No evident study on fiber properties | Usage of chemicals, incubation time duration | Yadav et al. (2017) |
| Bacillus cereus | Pectinase | Degumming of ramie and kenaf | 10 h bacterial treatment | Pectin hydrolysis selective medium | Weight loss was only 4% | Cheng et al. (2018) |
|
Bacillus licheniformis, Bacillus megatarium |
Pectin lyase | Flax degumming | Degumming was studied from 0h to 160h | Mixed system – advantageous, higher degumming rate | Weight loss was 6.33 and 8.56% | Ge et al. (2019) |
| Microbial consortium RAMCD407 | Pectinase | degumming process of ramie fiber | 56h degumming + 0.2% NaOH treatment | Ecofriendly, reuse of waste residues increases economic benefits | Prolonged incubation time | (Mao et al., 2019) |
| Bacillus cereus | Calcium alginate immobilized and crude Pectin lyase | degumming of plant fibers | 15 h, 45oC | Maximum galacturonide released | No tensile strength analysis | Kohli and Gupta (2019) |
3. Xylanase
Xylan, a polysaccharide, constitutes β-xylopyranose residues as spinal with glycosidic linkages. Xylan and pectic substances are inter-linked by xylan-glucan-protein complexes (Selvendran, 1985). The xylan was found to be aggregated with hemicellulose via hydrogen-bonding interactions. The xylanase enzyme system comprises the repertoire of hydrolytic enzymes which act cooperatively for the conversion of xylan into constituent sugars such as acetyl xylan esterase, β-xylosidase, α-L-arabinofuranosidase, α-glucuronidase, β-1,4-endoxylanase, and phenolic acid (ferulic and p-coumaric acid) esterase. Production of xylanase can be by bacteria, fungi, yeast, and marine algae. The inherent sources of xylanase were identified to be Bacillus among all the bacterial species. The other species of Bacillus were also reported in terms of considerable xylanolytic activity (Gupta et al., 2015). Trichoderma reesei LM-UC4 E fungus was mixed with Aspergillus niger ATCC 10864 and A. phoenicis QM 329 to produce xylanase using Solid-state fermentation (Rabello et al., 2014). The three-dimensional image of the xylanase enzyme is illustrated in Fig. 2.
 Fig. 2
Fig. 23.1. Production of xylanases by fermentation
Solid substrate fermentation is becoming an attractive process in xylanase enzyme production because of its higher productivity and lower synthesis cost. Using lignocellulosic materials in place of commercial xylan in the fermentation can deduct the synthesis cost of xylanase (Senthilkumar et al., 2005). The important factors which affect microbial xylanase synthesis are the usage of the transparent substrate and potent microbes. Utilizing bountiful and fruitful farming residues can enhance production and reduce the manufacturing cost in further applications. The various research group studied Xylanases production from different species by solid substrate fermentation and are listed in Table 4.
Table 4. Xylanases production by various species using different substrates.
| Species | Substrate | Type of Fermentation | pH | Temperature | References |
|---|---|---|---|---|---|
| Aspergillus niger DFR-5 | Mixed fruit peel waste | Solid substrate fermentation | 5.0 | 37 °C | Pal and Khanum (2010) |
| Bacillus arseniciselenatisDSM 15340 | Wheat bran | Solid substrate fermentation | 8.0. | 50 ◦C | Kamble and Jadhav (2012) |
| Aspergillus fumigatus SK1 | Palm oil waste | Solid substrate fermentation | 4.0 | 70 °C | Ang et al. (2013) |
| Mucorindicus, Mucorhiemalis, and Rhizopus oryzae | Wheat bran | Solid substrate fermentation | 4–5 | 40.0, 43.4, and 43.4 °C, respectively. | Behnam et al. (2016) |
| Aspergillus niger, Aspergillus oryzae, and Aspergillus awamori | Wheat bran | Solid substrate fermentation | 4.0 | Yin et al. (2018) | |
| Endophytic fungi | Lignocellulosic substrates | Solid substrate fermentation | 4.5–6 | 30–70 oC | Marques et al. (2018) |
| Aspergillus nigerCCUG33991 | waste of corn, bran | Solid substrate fermentation | 4–5 | 28–35 oC | Khanahmadi et al. (2018) |
| Trichoderma asperullum UC1 | Raw oil palm frond leaves | Solid substrate fermentation | 3.0 | 30 °C | Ezeilo et al. (2019) |
| Rhizopus oryzae UC2 | Raw oil frond leaves | Solid substrate fermentation | 3.0–5.0 | 50–60 oC | Ezeilo et al. (2020) |
| Aspergillus niger | Rice husk | Solid substrate fermentation | 5.0 | 30 °C | Afiqah Razali et al. (2020) |
| Aspergillus niger | Corn hub and saw dust | Solid substrate fermentation | 3.0 | 45 °C | Ire et al. (2021) |
3.2. Application of xylanases in degumming
Fibers can be degummed by using xylanolytic enzymes with pectinolytic enzymes (Puchart et al., 1999). Even though pectinases pursue a potent role in degumming, the involvement of xylanase is also considered (Cao et al., 1992). The conventional slow retting process can be replaced by enzymatic methods, creating a platform for new fiber liberation technology. The enzymatic degumming creates less damage to the cellulosic substance of the natural fiber. Xylanases play a major role in the degumming of ramie fibers. Extensive research has been carried out on the degumming of various natural fibers by Xylanaces produced from different species. The fibers with xylanase treatment were found to possess high linear density fibers with low prices compared to the Laccase treatment as it ends up with lower linear density and high cost. Some of the research findings are listed in Table 5.
Table 5. Degumming of natural fibers by xylanases using different species.
| Species | Enzyme | Application | Conditions | Observation | Problems /Disadvantage | References |
|---|---|---|---|---|---|---|
| Bacterial consortium | Xylanases pectinases | Screening of ramie-degumming strains | Diversity, characterization of degumming strains | Pectinase was the key enzyme of ramie degumming, | Some strains produced cellulase and degraded cellulose | Duan et al. (2012) |
| Bacillus sp. HG-28 |
Xylanases Pectinase |
Ramie fiber |
16h microbial degumming + NaOH treatment |
Efficient than chemical degumming, gum content reduction | Usage of chemicals, cellulase production | Fan et al. (2015) |
|
Bacillus cereus P05 Pseudomonas sp. X12 |
Xylanases pectinases | Ramie fiber | 60 h retting process | Breaking strength, gum removal increased | Prolonged retting time | Wang et al. (2017) |
| bacterial consortium RAMCD407 | Xylanases pectinases | ramie degumming | 56 h followed by 0.2% NaOH treatment. | Recovery of pectin crystals and xylan chips | Usage of chemicals, cellulase production | Mao et al. (2019) |
4. Laccase
Laccases belong to the oxidases family and are widely found in a diverse group of microorganisms. Production of Laccase from Monocillium indicum was demonstrated with staging activity by Johannes and Majcherczyk. (2000). The phenol-oxidase and ligninolytic property of laccase was synthesized by Pycnoporus cinnabarinus and Pycnoporus sanguineus, respectively. The three-dimensional image of the Laccase enzyme is illustrated below in Fig. 3.
 Fig. 3
Fig. 34.1. Production of Laccase by fermentation
Both solid-state and submerged fermentation methods produced the Laccase enzyme. Solid substrate fermentation has advantages over submerged fermentation in yield rate, waste production, lower energy needs, cost-effective media. Literature studies suggest solid-state fermentation has immense application in synthesizing antibiotics, biocides, surfactants, and ligninolytic enzymes by using agricultural residue as substrates. It can be adopted by inert solid support for easier recovery of extracellular products and is cost-effective (Rodríguez Couto and Sanromán, 2019). Many researchers have synthesized Laccase enzymes using different species and substrates and are listed in Table 6.
Table 6. Laccase enzyme production by various species and substrates.
| Species | Enzyme | Substrate | Fermentation | Reference |
|---|---|---|---|---|
| Trametes vesicolor | Crude laccase | Agro residues | Solid substrate fermentation | Stoilova et al. (2010) |
| Lenintu laedodas | Laccase and endoglucanase production | Reed grass, beanstalks, wheat straw | Solid substrate fermentation | Philippoussis et al. (2011) |
| Coriolopsis caperata RCK2011 | Laccase | Corn steep liquor | Solid substrate fermentation | Nandal et al. (2013) |
| Aspergillus species | Laccase | Winery and olive mill waste | Solid substrate fermentation | Salgado et al. (2014) |
| Pleurotus ostreatus | laccase | Sugarcane bagasse | Solid substrate fermentation | Karp et al. (2015) |
| Pseudolagarobasidium acaciicola LA1 | Laccase production | Parthenium biomass | Solid substrate fermentation | Adak et al. (2016) |
| Pleurotus ostreatus | laccase and peroxidase of and lignin | Potato peel waste | Solid substrate fermentation | Ozcirak and Ozturk (2017) |
| Pleurotus eryngii | Laccase | Peach waste | Solid substrate fermentation | Akpinar and Ozturk (2017) |
| Trametes versicolor IBL-04 | Alginate-chitosan Immobilized Laccase | corncobs | Solid substrate fermentation | Asgher et al. (2018) |
| Ganoderma lucidium | Laccase | Wood waste | Both fermentation | Rodrigues et al. (2019) |
| Trametes versicolor | Laccase | Tea residue | Solid substrate fermentation | Xu et al. (2020) |
| Galactomyces geotrichum | Laccase | Olive leaves | Solid substrate fermentation | Pourkhanali et al. (2020) |
| Halopiger aswanenesis | Lignolytic enzymes | Wheat bran | Solid substrate fermentation | Chauhan and Choudhury (2021) |
| Pycnoporus sanguineus | Laccase | – | Solid substrate fermentation | Hafid et al. (2021) |
4.2. Application of laccases in degumming
Laccase enzyme will deteriorate the dense, sticky material in the interior core and lateral surface, producing cellulose-rich fibers. Laccases are reaction-specific catalysts with focused performance, inexpensive, and environment-friendly, leading to the acceptance of laccase in degumming applications. Laccases are potently capable of lignin degradation, and therefore it can be accomplished in bast fibers degumming. The potential of laccase enzymes in refining bamboo fibers, enhanced lignin removal was successfully reported by Liu et al. (2012). They also highlighted bamboo fibers refining xylanase and laccase with limited loss of strength (Sreenivas et al., 2020). George et al. (2016) reported the fast growth of enzyme application in natural fiber modification. Some of the observations on degumming of fibers by Laccases are listed in Table 7.
Table 7. Degumming of fibers by Laccases.
| Process | Enzyme | Application | Observation | References |
|---|---|---|---|---|
| Laccase mediator system treatment | Laccase | Changes of lignin in the jute fiber | Improves mechanical properties, surface hydrophobicity | Zhang et al. (2014) |
| Enzymatic treatment | Laccase, Xylanase | Study on banana fiber properties | Lignin, hemicellulose removal was higher | Vishnu Vardhini and Murugan (2017) |
| Laccase and TEMPO mediated system | Laccase and 2,2’tetramethylpiperidine-N-oxyl | Grafting of silk fiber | The concentration of amino group decreased, self cross-linking of fibers | Zhou et al. (2017) |
| Chemo-enzymatic modification | Laccase | Hemp fiber degumming | Improves moisture absorption, water retention value, | Yeping et al. (2019) |
| Enzymatic treatment | Laccase, Pectinase | Banana fiber degumming | Removal of pectinolytic substances, surface smoothening of banana fiber | Paramasivam et al. (2020) |
5. Analysis of degummed natural fibers
Degummed natural fiber exhibits modification in the properties and composition of fibers. Efficient degumming results in the significant removal of hemicellulose substances. The hemicellulose removal can be confirmed by analyzing the chemical composition of natural fibers before and after degumming. In our study, we have illustrated the analytical results of degummed natural fibers. The analytical results included in our study are tensile analysis, lignin quantification, hemicellulose and cellulose content, density, moisture, and ash content.
5.1. Determination of tensile strength
Tensile strength measures the breaking strength, force resistance, and yield strength of the natural fibers. It can be analyzed using tensile identifying equipment under the ASTMD3822 standard. Microwave pretreatment was experimented with hemp fibers to remove the hemicellulose substance. The tensile strength of degummed hemp fiber was increased to 12.66 cN from 10.26 cN (Han et al., 2011). The Peroxide treatment of ramie fibers possesses a moderate decrease in the length, tenacity, and elongation property. The decreased tenacity corresponds to the breakage of microstructures and cellulosic fibrils (Li and Yu, 2014). Degumming banana fiber with the steam explosion method resulted in higher breaking strength of 156.37 Mpa (Sheng et al., 2014). An increase in the concentration of alkali in degumming of lotus fibers exhibits a decrease in the reduction of braking force from 5.31 cN to 2.37 cN and reduced elongation percentage (Li and Fu, 2015). The oxidized ramie fibers have increased tenacity to 6.18 cN/dtex (centinewton decitex) from 3.02 cN/dtex (centinewton decitex). The increase in tenacity is due to the successful removal of non-cellulosic substances, delignification of hemicellulose, and pectin degradation.
Further, the tensile property of fiber is directly proportional to the degree of polymerization. The degummed ramie fiber has an inclined polymerization degree. The increase in tenacity and polymerization degree of ramie fiber was due to the degradation of non-cellulosic chains to smaller compounds (Li et al., 2015). Meng et al. (2017) reported that the alkali coupled with magnesium hydroxide treated ramie fiber has resulted in the tenacity and elongation of ramie fiber. The substitution rate of magnesium hydroxide above 20% in the degumming process causes a decrease in the tensile strength of the ramie fiber. Enzymatic treatment assists the removal of fragile structures and modifies the molecular behavior of fibers. The modification in molecular behavior corresponds to the amplified tensile strength compared to the chemical degumming methods (Balakrishnan et al., 2019).
5.2. Determination of lignin, hemicellulose, and cellulose content
The major parameters that influence the chemical composition of fibers are the locality of the plant, maturity, degumming methods, and characteristics of the fiber. The matrix formation of cellulose and hemicellulose was due to the presence of non-cellulosic layers. The lignin content corresponds to the rigidity of the plant. Li and Yu. (2014) reported that the per-oxide treatment of ramie fibers has successfully reduced the hemicellulose content from 14% to 5%. But the peroxide treatment was less efficient in lignin removal of ramie fibers. The oxidized ramie fibers have increased cellulose content from 73% to 95% and decreased non-cellulosic content from 13% to 3%. The degree of gummy substance removal is reflected in the reduction of residual gum content. The efficient pectin and non-cellulosic substance removal lead to finer and separated cellulosic fibers (Li et al., 2015). The peracetic acid-treated jute fiber has increased cellulose content, reduced hemicellulose, and lignin content. The reaction of the lignin with peracetic acid causes electrophilic hydroxylation of aromatic rings, oxygenated groups. The hydroxylation results in the formation of quinone, lactone, and muconic acid and demethoxylation (Duan et al., 2017). Jiang et al. (2018) demonstrated that the green degumming of ramie fibers results in the efficient removal of the gummy substance from 27% to 5%. Further, green degumming has improved the fineness of ramie fiber to 1400Nm and also increased the whiteness of fiber to 50%. The degumming of kenaf fiber has significantly removed hemicellulose and lignin content. Zhang et al. (2019) observed that the novel degumming of kenaf fiber has also increased the percentage of cellulose content and absorption rate.
5.3. Determination of density and diameter
ASTM D 1505-03 is the standard method to measure the density of the fibers. Linear density can be calculated using ASTM D 1577-07. Solvents used for the determination of density are xylene and carbon tetrachloride. Fiber fineness can be measured in Tex (weight in grams per unit length of fiber). The efficient removal of hemicellulose substance corresponds to an increase in linear density of fiber and finer fibril molecules (Devireddy et al., 2018).
5.4. Determination of moisture and ash content
The moisture of natural fiber will be measured and expressed as a percentage. The oven conditions the fiber samples at 105oC, and the weight (WI) will be determined before and after conditioning as per ASTM D 2495-07 standard. The ash content determines the inorganic residues after heating at 600oC for 4–5 h. The banana fibers were subjected to the steam explosion at 800oC. The ash content of banana fiber was reduced to 1.72 ± 0.12 from 7.02 ± 0.32 by degumming the banana fiber with a steam explosion (Sheng et al., 2014). Li and Yu (2014) reported that the ramie fibers were degummed by alkali, peroxide, and isopropyl alcohol coupled with peroxide and compared with control ramie fibers. The ash content of control, alkali, peroxide, peroxide associated with isopropyl alcohol were 2.45%, 1.42%, 1.02%, and 1.24%, respectively(Li and Yu, 2014). The oxidation-reduction potential on the degumming of ramie fibers with hydrogen peroxide has decreased the ash content of ramie fibers from 2.45% to 0.28% (Li and Yu, 2015). Similarly, Song et al. (2019) reported that the combined steam explosion and chemical degumming of kudzu fiber had reduced the ash content.
5.5. Weight loss analysis
Weight loss analysis is done to identify the gum loss of control and degummed natural fibers. The weight loss of degummed fibers can be calculated concerning the initial weight of the fibers. Many researchers highlighted the advantages of enzymatic degumming in significant weight loss. Ibrahim et al. (2019) reported that the scouring of cotton fabrics with polygalacturonase enzyme resulted in 5.2% weight loss (Ibrahim et al., 2019). The gum content was reduced to 11.6% from 25.9% on enzymatic degumming of ramie fiber (Jiang et al., 2018). The literature studies show that enzymatic treatment of jute fiber contributes to a weight loss of 12–17%. Similarly, the cotton fabric treated with enzymes also resulted in a weight loss of 10% (Rajulapati et al., 2020). The weight loss percentage of enzymatic degummed fiber was significant compared to conventional degumming methods.
6. Characterization of degummed fibers
Characterization studies can examine the surface modification of fibers. The characterization studies reported in this research include Microscopy analysis, SEM, FESEM, ESEM, FTIR, XRD, NMR, and thermal analysis. Characterization studies will examine degummed fibers to identify the pros and cons of degumming methods.
6.1. Microscopy analysis
An optical microscope, commonly known as a Light microscope, is used to visualize the magnified view of the sample through the series of lens magnification under visible light. Polarized light microscopy is one of the members of the optical microscope by involving the illumination of the sample under polarized light. A fluorescence microscope is also a member of the optical microscope, which employs not only fluorescence but also few features such as scattering, reflection, attenuation, and absorption. Microbial retting of ramie fibers exhibits an average diameter of 32um, greater than chemically treated rame fiber (Angelini et al., 2015). The mechanically extracted banana fiber was comparatively rough and less significant under the light microscope (Balakrishnan et al., 2019). For a light microscopic view of ramie fibers, bast tissue, gum debris, and treated ramie fiber were placed on a microscopic slide with the aid of tweezers and a drop of distilled water. For a Polarized microscopic view of ramie fibers, samples were illuminated with a mercury lamp for 212 milliseconds. The reference of ramie xylan, pectin, and lignin under illumination was an indigo blue image, dark orange, and red image, respectively. The replacement of reference color with bright white infers the removal of non-cellulosic substances (Mao et al., 2019). The Autofluorescence signal of the non-cellulosic ramie gum component is visualized by the dark orange fluorescent signal emitted by ramie pectin particles. Similarly, the ramie xylan particles emit bright indigo fluorescent signals, and lignin emit a red fluorescent signal.
6.2. Morphological surface studies
Scanning Electron Microscopy generates signal diversification at the outer layer of the solid specimen by high-energy electrons with a focused beam. Generated signals produce a 2-dimensional image that reveals knowledge of sample texture and chemical composition. Comparative analysis of the surface of sample fibers with control can be examined using a scanning electron microscope. Removal of gum material from the exterior of the bast fiber cell and their modification into single bast fibers can be examined by SEM image (Beltran et al., 2002). Li and Fu (2015) demonstrated that the alkali degumming of lotus fibers results in the removal of binding agents without damaging the nature of the fibers. Oxidation degumming of ramie fiber has resulted in fine pits and plaques, thereby losing fibrous structures (Li et al., 2015). Duan et al., 2017 reported that the SEM image of acid-treated Jute fiber had shown the partial removal of gummy substance with few elementary fibrils in the primary layer of jute fiber. Researchers also reported that most gummy substances were detached from the exterior of the ramie fiber bundle by microbial degumming of ramie fibers. The SEM images of degummed ramie fiber surface were clear and smooth (Mao et al., 2019). Similarly, research on degummed kenaf fiber also resulted in reduced gum materials compared to raw kenaf fibers. Further, literature studies also infer that green degumming, such as microbial and enzymatic degumming of ramie fiber, had comparable efficiency to the conventional fiber degumming analysis (Jiang et al., 2018).
6.3. Field emission scanning electron microscope
The field emission scanning electron microscopy (FESEM) is employed to visualize the topographic details of a sample surface, fractionated objects, nanomaterials, and nanocomposites. FESEM is one of the types of electron microscope which scans the sample surface with a high-energy beam of electrons in a raster scan pattern. Rajulapati et al. (2020) reported that the FESEM image of microbially degummed jute fibers, chemical degummed jute fibers, and control jute fibers were compared. The microbially degummed jute fibers exhibit light brown color, smoother surface, removal of pectin and wax. Whereas, chemically treated jute fibers and control fibers were in yellow and dark brown color respectively. Similarly scouring of cotton fabrics was also analyzed by FESEM. The image of enzymatically treated cotton fabrics, chemically treated cotton fabrics, and control fabrics were light yellowish-white, whitish, and yellowish-white. The smoothness on the surface of enzyme-treated cotton fabrics was efficient compared to other methods(Rajulapati et al., 2020).
6.4. Environmental scanning electron microscopy
Environmental scanning electron microscopy (ESEM) is a unique technique that involves the examination of uncoated biological material and industrial materials with a high electron beam in a high chamber pressure atmosphere of water vapor. The major applications of ESEM are dynamic experiments, drying, and crystallization. ESEM employs a scanned electron beam, electromagnetic lenses for the electron beam's direction on the specimen's surface. The interaction of specimen surface and electron beam results in various signals collected with appropriate detectors. The ESEM micrographs of electrospun silk fibers were circular in cross-section and smooth surface. The electrospun fibers were spun with a 3 kV/cm (Ayutsede et al., 2005). Li and Fu (2015) reported that the lotus fibers plucked at different growth stages were examined by ESEM analysis. The surface of the control lotus fibers was coarse, connected by pectin to form fiber bundles. The surface of degummed lotus fibers was individual in longitudinal view with the removal of gummy substances. The ESEM image of enzymatically treated flax fibers showed significant sticky substance removal, leading to the formation of a single fiber (Xiang et al., 2020).
6.5. Functional group analysis
The physiochemical properties of the lingo cellulosic materials can be investigated by a non-destructive method. FTIR analysis can be executed to examine the modifications in the spectral regions of bast fiber structures before and after treatment (Beltran et al., 2002). The intensification of cellulose composition can be observed by the sharpening peaks attributed to the fibers' crystallinity (Cherian et al., 2008). The height at 1,735cm-1 corresponds to the carboxylic esters in pectin and wax. Lignin usually peaks at 1,510 cm-1 which disappears on degumming of Hemp fibers (Han et al., 2011). Microwave-assisted degumming of hemp fibers exhibits the disappearance of the lignin region, unchanged cellulose region, and presence of glycosidic bonds (Han et al., 2011). The spectrum of peroxide treated ramie fiber exhibits higher C–H stretching, CH2 symmetric bending, and C–O stretching intensities. The higher intensities correspond to the successful removal of gummy substances (Li and Yu, 2014). The oxidation degumming of ramie fibers causes breakage in the hydrogen bond structures.
Further, increased oxidation rate causes increased stretching vibrations, thereby leading to aldehyde and carboxyl groups (Li et al., 2015). The alkali degummed ramie fiber research highlighted the efficiency of magnesium hydroxide in degumming relies on the substitution rate. The substitution of magnesium hydroxide above 20% affects the cellulosic nature of the fibers (Meng et al., 2017). Degummed jute fiber was found to have reduced peak at 2,900 cm-1 and 1,420 cm-1 indicating the removal of hemicellulose and lignin (Duan et al., 2017). FT-IRM analysis of degummed kenaf fiber shows the breakage of hemicellulose substance, making the surface of the kenaf fiber smooth and clean. The researcher also highlighted that microbial degumming was more efficient in removing a non-cellulosic substance from the kenaf fiber (Zhang et al., 2019). Green degumming of ramie fiber effectively removed the lignin in degummed ramie fibers (Jiang et al., 2018). The alkali degumming of lotus fibers disturbs the hydrogen bond of cellulose and also cause substitution of sodium atom in place of hydrogen. The ATR-FTIR analysis of enzymatic degummed jute fibers possesses significant change in alkane and alcohol groups of the pectin and also loss of methylation groups in the pectin region (Rajulapati et al., 2020)